

分形是现代数学和非线性科学研究领域中一个十分活跃的分支。由于世界的本质是非线性的,因此,分形的应用领域非常广泛,是当前研究的一个热点。而分形生长是分形中重要的研究课题之一,从煤灰微粒的凝聚、闪电的形态、癌细胞的扩散与凝聚,到自然界中植物的生长等复杂而又不规则的实际聚集生长现象,都是分形生长的实际案例。因此,分形生长的理论研究与应用分析具有重要的理论意义和实际应用价值。然而,实际环境中的聚集生长过程总是会受到像磁场、温度等许多复杂因素的影响,从而使得生长粒子的形态变化多端,或者使得生长粒子的生长速度爆发性增长,以至于给自然界和人类社会带来很多负面的影响。因此,实际环境中的聚集生长形态的预测和控制,以及生长速度的控制具有重要的现实意义。
1981年美国密捷安大学Witten和Sander首次提出了扩散限制凝聚(diffusion limited aggregation)的分形生长模型(简称DLA模型)[1], 该模型用来描述像悬浮在大气中的煤灰、金属粉末或烟尘的扩散过程。近几年来对分形生长的研究主要体现在2个方面:一是生长模型改进[2-5];二是分形生长的应用[6-8]。针对目前国内外分形生长控制研究热点,本文将从定量分析的角度将控制的思想和方法引入到分形理论中,研究对实际环境中的分形生长的形态变化进行有效的控制。基于已有DLA模型生长速度慢的问题,应用多粒子发射理论,提出一个改进模型——多粒子扩散限制凝聚(multi-particle diffusion limited aggrega-tion)模型,简称MDLA模型,探索和研究该模型下分形生长的形态控制和模拟问题,使其产生出实际应用中需要的控制效果。
1 MDLA生长模型的提出标准DLA模型中粒子是从远离中心的某个点逐个发射的, 当实际应用中需要生成一个由成千上万个粒子生成的大的团簇时, 再用此方法发射粒子, 时间效率就很低下。为了加快粒子凝聚速度, 本文从改变粒子发射方式和粒子死亡条件的角度出发, 提出改进模型-MDLA生长模型, 该模型与标准DLA模型有3个区别:
1) 粒子不是逐个从矩形边缘发射的, 而是被全部初始化;
2) 粒子运动方向由4个方向变为8个方向;
3) 粒子消失的条件设定为与其初始位置的距离大于某个常数。
MDLA模型的生长规则是由迭代产生的, 所以具有分形的自相似性, 同时MDLA模型还具有屏蔽效应和非整数性。MDLA模型定义了8个方向, 即上、下、左、右、左上、左下、右上、右下, 粒子向8个方向运动是等可能的, 每个方向概率均为1/8。
MDLA算法是一次性初始化多个粒子, 被初始化后的粒子都在做随机布朗运动。只要某个粒子满足与团簇粘贴的条件, 该粒子(候选点)就会成为生长簇的一部分, 停止运动。因此, 候选点的选择不唯一, 同时生长点的选择也是随机且不唯一, 只要有粒子能够与之粘贴, 该粒子就是生长点。因此, 在MDLA模型中, 同一时刻生长点和候选点的选择都不是唯一的。
实验结果表明, MDLA算法要求n个寄存器, 记录粒子的当前状态, 包括位置、颜色、大小、半径。时间复杂度接近O(n), 以空间换取时间, 从而加快粒子的凝聚。
因MDLA模型只是改变了粒子初始化方式, 故也与标准DLA模型一样满足Laplace方程。求解Laplace方程的数值解是相当困难的, 尤其当方程的阶数比较大时。因此运用有限差分理论, 并结合第一边界条件, 把定解问题转化为求未知函数u(x, y)在节点上数值为未知量的线性方程组的解的问题。定义A为系数矩阵, 与网格节点的划分和编号方式有关, b为方程右端的常数向量, 由边界条件确定, 因此系统方程转化为如下线性方程组
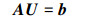
|
式中,U是扩散方程的解。
在平面上, u是x和y的函数, 不带源项的扩散系统方程可以写成

|
将平面网格化, 令

|
式中, (x0, y0)是xoy平面上的一个随机点, h, τ分别表示x, y方向步长。由于
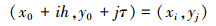
|
故(xi, yj)=(i, j), (x0, y0)=(0, 0)。
当h=τ时, 只要粒子数目确定, 粒子随机向周围8个方向运动一步, MDLA生长基本上是一致的。当h=τ=1时, 就是各个方向的迭代步长为1的MDLA(云团多粒子凝聚扩散)。
当h≠τ即x, y方向步长不一致时, 理论上是哪个方向的步长大, 粒子在哪个方向上运动的范围就广, 粘贴上的概率就大, 最终生长的形状应该是接近平行于该方向, 表明相同生长条件下, 粒子凝聚更快。
在MDLA模型中, U代表粒子的粘贴概率矩阵, 当U值全是1, 此时没有任何干扰项, 即扩散方程的右端项为0, 此时初始化的粒子向各个方向生长的可能性是一样的。这样, 粒子在运动过程中, 只要满足粘贴条件, 该粒子就会成为生长簇的一部分。
2 迭代步长对生长形态的影响当粒子同时满足下面3个条件时, 粒子可能会进行更新, 继续走下一步:
1) 粒子没有粘贴在团簇上;
2) 粒子当前位置与其初始位置的距离小于一个常数;
3) 粒子还没有逃出窗口边界。
因此, 粒子运动步长的改变, 使粒子相对容易走到聚集团旁边或离开凝聚区域, 进而决定粒子的运动方向, 影响最终生长簇的生长形态。
做出以下推测:粒子迭代步长的增大, 使粒子更快凝聚或更快停止生长(溢出边界条件或与其最初位置的距离超过一定值)。特别当x, y方向的步长不一致时, 分形图形状会发生改变。同时,迭代步长的变化可能会影响分形维数。
图 1表明了相同初始化条件下, 粒子在运动的过程中, 采用不同迭代步长d得到的单种子生长簇。

|
| 图 1 不同迭代步长的生长簇 |
表 1列出了对应图 1a)~1c)的生长簇的回旋半径r[9]、粒子数n、分形维数Df的变化值。表 1说明不同步长生长簇的粒子数和分形维数基本不变, 变化的是回旋半径。由此可知, 变化的步长在一定程度上会影响生长簇的局部特性, 而分形维数并未受到影响。
在MDLA模型中, 规定粒子向其周围8个方向随机运动, 假定M(x), M(y)分别为x, y坐标方向的搜索步长。搜索步长是粒子在其周围搜索是否存在生长点的一种衡量尺度, 步长越大, 越有利于生长。图 2表明了不同坐标方向的不同搜索步长的生长图。图 2a)表明, 当x方向的搜索步长与y方向的步长时相等时, 粒子均匀生长。当x方向的搜索步长大于y方向的搜索步长时, 粒子有向x方向粘贴的趋势, 最终长成侧向“V”字形(见图 2b)), 而且某个方向的搜索步长越小, 生长簇的分叉越小, 方向步长的改变直接影响生长簇的生长形态。

|
| 图 2 不同方向不同搜索步长M的生长图 |
粒子半径代表着粒子大小, 粒子在运动过程中可以认为是恒定不变的, 默认条件下粒子半径是1, 这样当粒子运动步长为1时, 相当于走粒子半径大小的距离。那么当粒子运动步长不变时, 粒子半径对生长簇的影响是什么呢?下面从实验角度探讨半径R的作用。
当粒子半径R=1.0, 2.0, 3.0, 其他条件均相同时生长图如图 3所示。实验结果表明, 粒子半径越大, 生长簇的颜色越深, 生长簇看起来越密集, 但生长簇上的粒子数并没有变很大, 同时发现分形维数基本保持在1.75左右。由此可以得出, 粒子大小影响的是生长簇的局部性质, 而对全局性质影响不大, 这从分形维数的变化可以看出来。

|
| 图 3 粒子半径不同的生长簇 |
下面从改变生长参数入手, 如方向概率、粒子初始化位置、粘贴位置、种子数目等, 探讨生长簇的形态控制问题。
图 4模拟的是初始化位置不同, 不同形态的生长簇。图中点密集地方代表初始化的粒子, 稀疏且出现分叉的点代表生长簇上的点, 由图 4可知, 生长簇的最终形态与初始化状态有关。

|
| 图 4 不同初始化方式得到的生长图 |
下面探究方向概率对生长形态的影响。在MDLA模型中, 方向概率是指粒子在任意时刻, 任意位置, 往某个方向运动的可能性大小。在此模型中方向概率有8个方向, 所以粒子朝向某个方向运动的概率是相等的, 均为1/8, 如果某个方向的方向概率为1说明粒子仅向该方向运动。图 5表明了仅方向概率不同时的生长图。图 5a)中粒子向上的生长概率为1, 表示粒子只向上运动, 所以粒子的粘贴主要在种子下方, 形成一个倒立的“锥形”。而图 5b)是各方向的方向概率共同作用的结果, 哪个方向的概率大, 粒子朝向哪个方向运动的可能性就大, 从而定性决定生长簇的大致形态。图 5a)和图 5b)中的生长图方向概率不相同,分形维数Df也不相同。

|
| 图 5 方向概率不同得到不同维数Df的生长图 |
粒子粘贴条件是粒子能粘贴到生长簇上的条件。首先在整个窗口中初始化所有粒子, 所有粒子做随机布朗运动, 生长簇上的粒子除了满足基本粘贴条件外, 还应满足被预先设定的粘贴位置条件。图 6表明了粒子粘贴位置不同而产生的不同的生长簇, 生长簇的粒子横坐标x, 纵坐标y满足一定的函数关系。

|
| 图 6 粒子粘贴位置不同的生长簇 |
另外初始化种子数不同,可导致同一时刻可以有多个粒子与团簇发生碰撞,最终表现为生长形态的差异。同时由于分形的非整数性及自相似性,可以推断出其分形维数也不同。图 7表明了不同的种子个数的生长簇。

|
| 图 7 不同种子数的生长簇 |
本文基于已有DLA模型,提出了一个改进模型——MDLA模型。从改变模型参数入手,通过分析、观察、对比等手段,研究生长形态的定向控制问题,并用仿真实例表明了控制方法的有效性,为进一步研究分形生长的定向控制奠定了基础。
| [1] | SANDER L M, WITTEN T A. Difusion-Limited Aggregation:a Kinematic Critical Phenomenon[J]. Physical Review Letters, 1981, 47: 1400-1403. DOI:10.1103/PhysRevLett.47.1400 |
| [2] | BRAGA F L, RIBEIRO M S. Diffusion Limited Aggregation:Algorithm Optimization Revisited[J]. Computer Physics Communications, 2011, 182(8): 1602-1605. DOI:10.1016/j.cpc.2011.04.005 |
| [3] | KASPER R K, LILIAN D M, RUUD VAN OMMEN J, et al. Optimizing Off-Lattice Diffusion Limited Aggregation[J]. Computer Physics Communications, 2014, 185(3): 841-846. DOI:10.1016/j.cpc.2013.12.003 |
| [4] | BRAGA F L, MATTOS O A, AMORIN V S, et al. Diffusion Limited Aggregation of Particles with Different Sizes:Fractal Dimension Change by Anisotropic Growth[J]. Physica A, 2015, 429: 28-34. DOI:10.1016/j.physa.2015.02.050 |
| [5] | GHOSH A, BATABYAL R, DAS G P, et al. An Extended Fractal Growth Regime in the Diffusion Limited Aggregation Including Edge Diffusion[J]. AIP Advances, 2016, 6(1): 1-2. |
| [6] | KATYAL N, BANERJEE V, PURI S. Fractal Signatures in Analogs of Interplanetary Dust Particles[J]. Journal of Quantitative Spectroscopy and Radiative Transfer, 2014, 146: 290-294. DOI:10.1016/j.jqsrt.2014.01.017 |
| [7] | RISLER T. Focus on the Physics of Cancer[J]. New Journal of Physics, 2015, 17(5): 1-13. |
| [8] | MAJUMDAR S, TRUJILLO-REYES J, HERNANDEZ-VIEZCAS J A, et al. Cerium Biomagnification in a Terrestrial Food Chain:Influence of Particle Size and Growth Stage[J]. Environmental Science & Technology, 2017, 50(13): 6782-6792. |
| [9] | MEAKIN P. Diffusion-Controlled Cluster Formation in 2-6-Dimensional Space[J]. Physical Review A, 1983, 27(3): 1495-1507. DOI:10.1103/PhysRevA.27.1495 |
